
Puntos clave
- Muy pocos deportes usan sólo la resistencia o la fuerza. Prácticamente todos los deportes requieren una combinación de resistencia y fuerza.
- Resistencia y fuerza se pueden desarrollar simultáneamente en algún grado. Sin embargo, como la frecuencia y la intensidad de los aumentos de entrenamiento de resistencia, puede afectar al el desarrollo o mantenimiento de la masa muscular y la fuerza esta por conocer. Esta interacción entre la resistencia y la fuerza en la fisiología del entrenamiento se le conoce como, efecto de interferencia.
- El aumento de la fuerza y la masa muscular requiere la activación de la diana mamífera de rapamicina (mTOR). En conclusión, la molécula de señalización mTOR se activa al máximo por levantar objetos pesados y el consumo de comidas proteícas ricas en leucina.
- Las adaptaciones de resistencia ocurren cuando el estrés metabólico es más alto (el suministro de energía es bajo y la demanda de energía es alta). El estrés metabólico activa el monofosfato de adenosina activada por proteína quinasa (AMPK) y la nicotinamida adenina dinucleótido (NAD) dependiente de la (SIRT1).
- El efecto de entrenamiento concurrente se puede explicar a nivel molecular, por el hecho de que el estrés metabólico (aumento de la AMPK y SIRT1 ) pueden inhibir la activación de mTOR y la hipertrofia muscular.
- Hay que comprender que: a) la importancia de mTOR en el desarrollo de la masa muscular y la fuerza; b) el espacio temporal de la activación de mTOR; y c) el papel de la nutrición en la mTOR / activación / SIRT1 AMPK, programación del entrenamiento y plan nutricional puede ser desarrollado para maximizar la fuerza y la resistencia.
El entrenamiento concurrente en la fisiología del entrenamiento se le conoce como (la combinación de ejercicios con carácter aeróbico junto con tipo anaeróbico, o viceversa) es una práctica común para los atletas que buscan maximizar la fuerza y la resistencia, (como en Atletismo, Gimnasia deportiva, o la practica de Crossfit, que no es nada nuevo por cierto…). Hace más de 20 años, se observó por primera vez que la realización de ejercicio de resistencia (edurance) después de ejercicio de resistido (fuerza) podría tener efectos perjudiciales sobre las ganancias de fuerza, a través de ciertas interferencias (también llamado el fenómeno de interferencia). A nivel celular, los hidratos de carbono, y ciertas proteínas específicas se han sugerido para mediar esta interferencia creada en el entrenamiento concurrente; Sin embargo, en la actualidad, la razón fisiológica que hay detrás del efecto de entrenamiento concurrente permanece en gran medida desconocida. Mucho menos se sabe acerca de las estrategias nutricionales óptimas para apoyar la formación concurrente, pues por esta razón se necesitan enfoques nutricionales únicas o especiales. En esta revisión, voy a discutir la importancia de una correcta alimentación como la administración de suplementos de deportivos, usados con cabeza y conocimiento, tanto para la adaptación de resistencia (edurance, carácter aeróbico) como para el entrenamiento resistido (fuerza, carácter anaeróbico) y destacar las estrategias nutricionales adicionales que pueden beneficiar la formación concurrente. Por último, vamos a tratar de crear sinergia actual en la comprensión de la interacción entre las respuestas fisiológicas y enfoques nutricionales respecto a las recomendaciones prácticas para el entrenamiento concurrente

Introducción
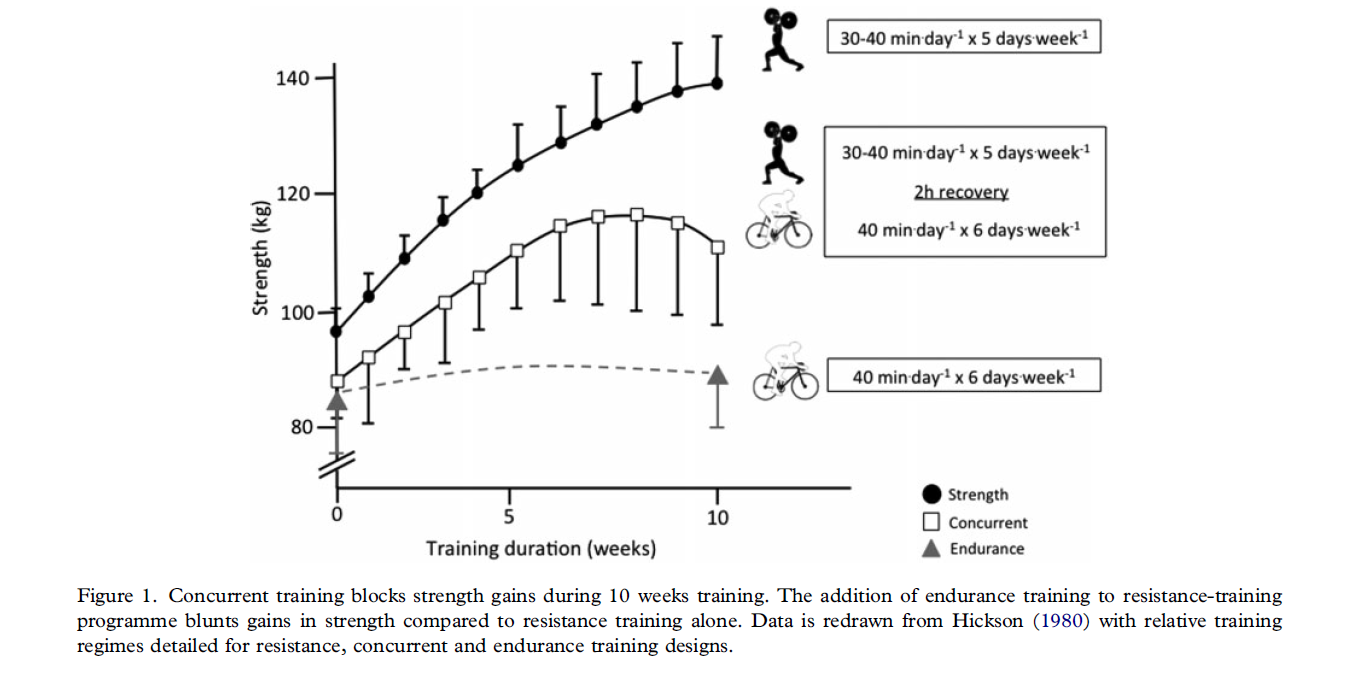
El fenómeno de interferencia producido por el entrenamiento concurrente, fue ideado por primera vez por Robert C. Hickson (Hickson, 1980). El utilizo un protocolo de alta intensidad, compuesto por ciclismo, y running, de carácter edurance, es decir un programa con componente concurrente. (Combinación de ejercicio resistido y edurance) en el mismo día de entrenamiento. Hickson demostró que las ganancias de fuerza con el añadido de ejercitico edurance era inferiores a las obtenidas en el protocolo el cual solo se hacia el ejercicio resistido de alta intensidad (Hickson, 1980). Estos datos demuestran por primera vez que el ejercicio de alta intensidad junto con edurance interfería con las adaptaciones y respuestas al entrenamiento de fuerza (figure 1). Numerosos estudios, investigan los mecanismos los cuales son responsables de (Fyfe, Bishop, & Stepto, 2014; Hamilton & Philp, 2013). Obviamente la nutrición es un rol enormemente importante en el entrenamiento concurrente, ya que se pueden llevar a cabo diferentes estrategia snutricionales como enfoque eficaz para compensar esa reducción de la fuerza ivestigada por Hickson (1980) y otros como (Ronnestad, Hansen, & Raastad, 2012).
Esta revisión que os hago, tiene como objetivo:
- Explicar brevemente los procesos moleculares y el funcionamiento del músculo esquelético como la adaptación al ejercicio de resistencia o resistido (fuerza), al ejercicio edurance como al ejercicio concurrente (ambos)
- Discutir diferentes estrategias nutricionales para mejorar la recuperación, o como obtener mayor adaptación del ejercicio físico, recordar a mayor adaptación, mayor progreso y rendimiento físico.
- Intento formular estrategias nutricionales atípicas, que podrían ser adecuados para el apoyo del entrenamiento concurrente.
Regulación molecular en las adaptaciones del ejercicio
El ejercicio edurance se basa en gran medida en la capacidad oxidativa del musculo esquelético, que a su vez es muy dependiente de la función de las mitocondrias como del suministro de sangre (Egan & Zierath, 2013). Por consiguiente el entrenamiento edurance aumenta la capacidad aeróbica gracias a la síntesis de la Adenosín Trifosfato (ATP) por el incremento de diferentes expresiones genéticas y regulaciones bilógicas, procesos mitocondriales, como la biogénesis y la angiogénesis (Egan & Zierath, 2013). El metabolismo del músculo esquelético y los mecánismos de estrés metabólico inducido por el ejercicio de resistencia se postula que crea cierta señal de caminos diferentes en el control de estas adaptaciones (Egan & Zierath, 2013). Está bien claro y establecido, que tras el inicio de la contracción, hay un rápido flujo, transitorio de sustratos, metabolitos y nucleótidos en el músculo esquelético, como Ca2 +, monofosfato de adenosina (AMP) y nicotinamida adenina dinucleótido (NAD +) (Egan y Zierath, 2013). Desde una perspectiva de adaptación (a mayor adaptación, mayor desempeño deportivo), esto es extremadamente importante, ya que muchos de éstos moléculas de señalización han sido identificadas como activadores clave, que en última instancia mejorarían la respuesta adaptativa.
El estrés metabólico conduce a la adaptación del músculo esquelético
Entre la amplia gama de proteínas reguladas por el ejercicio de resistencia, la AMP-activated proteína quinasa (AMPK) es probablemente la más estudiado en el contexto de la remodelación del músculo esquelético. La relevancia de esta proteína quinasa, se basa en su capacidad para detectar tanto el estrés metabólico como el almacenamiento de energía (Hardie, Ross, y Hawley, 2012). Todos deberíamos saber que el ejercicio físico activa el músculo esquelético AMPK en una manera dependiente de la intensidad, es decir, con el ejercicio a altas intensidades (por lo tanto una mayor rotación de ATP) resultando en una mayor activación de AMPK (Wojtaszewski, Nielsen, Hansen, Richter, y Kiens, 2000). Además del ejercico físico, es activada en situaciones de restricción de energía (por ejemplo, personas que están en déficit calórico con objetivo de pérdida de peso) o con el agotamiento de glucógeno en el músculo esquelético humano, sin embargo la actividad de la AMPK mejora tanto en condiciones basales como después del ejercicio agudo (Philp et al, 2013.; Wojtaszewski et al., 2003). A largo plazo, este proceso de activación de AMPK tras el ejercicio de resistencia puede aumentar capacidad oxidativa en el musculo esqueletico a través de la modulación de diferentes factores de transcripción y co-reguladores (Egan y Zierath, 2013); factor 1, factor nuclear respiratorio 2, factor mitocondrial, factor de transcripción y el co-activador de peroxisomas proliferativa ,como también el coactivador 1 alfa (PGC-1α), los cuales son centrales son moduladores de la biogénesis mitocondrial (García- Roves, Osler, Holmstrom, y Zierath, 2008; rockl et al., 2007). Los efectos positivos de la AMPK en la función mitocondrial son PGC-1α dependiente, que puede ser fosforilada y activada por AMPK in vitro (Jager, Handschin, St-Pierre, y Spiegelman, 2007), lo que indica una interacción dinámica entre las dos proteínas (Lerin et al, 2006;. Rodgers et al,. 2005). Este equilibrio añade otra manera por el cual el metabolismo del músculo esquelético puede ser modulado por el estado de la energía, ya que la actividad de SIRT1 es eficientemente inducida por un aumento en citosólica [NAD +] y la actividad GCN5 se cree que es regulada por el aumento de la disponibilidad de acetil-CoA (Philp y Schenk, 2013).
Además de las alteraciones en el ambiente metabólico, otras señales inducidas por la contracción del músculo esquelético también se han demostrado ser una importante señal de control de adaptación en el ejercicio, con el p38 activado por mitógenos de la proteína quinasa (MAPK) y el calcio dependiente de (CaMKII) y cAMP (PKA-CREB). Esta señalización se cree que desempeñan un papel clave (Egan y Zierath, 2013). Sin embargo, el cómo, estas proteínas podrían afectar a las adaptaciones de la fuerza aún no se ha demostrado con evidencia.
Regulación molecular de la resistencia en la hipertrofia del músculo esquelético
En contraste con el ejercicio de edurance, adaptaciones a ejercicio de resistido (fuerza) crónico requiere la activación de procesos anabólicos que en última instancia, promuevan una mayor acreción de proteínas, el aumento de la sección transversal del músculo esquelético área (CSA, es decir aumento del volumen muscular) como de la fuerza máxima (Egan y Zierath, 2013). En el músculo esquelético, la carga mecánica, promueve en estas adaptaciones principalmente a través de un aumento de la tasa de síntesis de proteínas y la biogénesis (Moore, Robinson, et al., 2009; Tipton, Ferrando, Phillips, Doyle, y Wolfe, 1999; Witard et al., 2014).
(Philips y metabolismo de las proteínas) La activación de la (mTOR) complejo 1 (mTORC1) parece ser un paso obligatorio y un punto de convergencia de las vías sensoriales en la carga mecánica (Philp, Hamilton, y Baar, 2011). La sobrecarga muscular se ha postulado que aumenta la actividad de la mTORC1 mediante el aumento de calcio citosólico [Ca2 +], ácido fosfatídico y leucina, además de la activación de la creatina quinasa extracelular y diacilglicerol quinasa (Miyazaki, McCarthy, Fedele, y Esser, 2011; Philp et al., 2011; Usted et al., 2014). La activación mTORC1 por los aminoácidos, en especial por la Leucina (enlace de metabolismo proteico), por otra parte, implica la activación de las GTPasas que median la translocación de mTORC1 desde el citosol a la superficie lisosoma (Shimobayashi & Hall, 2014). Puesto que los dos mecanismos son diferentes, la administración de aminoácidos esenciales potencia los efectos y adaptaciones del ejercicio resistido (fuerza) sobre la actividad mTORC1 en el músculo esquelético humano (Kakigi et al., 2014). Tras su activación, la proteina mTORC1 impulsa la sistensis a través de dos mecanismos principales (Figura 2):

(1) la fosforilación directa de la traducción eucariótica del factor de iniciación 4E (eIF4E) proteína de unión 1 (4E-BP1) y (2) la quinasa S6 1 (S6K1) (Shimobayashi & Hall, 2014). 4E-BP1 ejerce un efecto inhibitorio que libera eIF4E, promoviendo así la iniciación de la traducción dependiente, mientras que la fosforilación de S6K1 aumenta su actividad y la regulación de sus objetivos de abajo controlando la traducción, incluyendo S6 y eIF4B (Shimobayashi & Hall, 2014). El aumento de la síntesis de proteína inducida por el ejercicio de resistido en humanos en el músculo esquelético se ha demostrado que coincide con los niveles más altos de 4E-BP1 y S6K1, aunque los informes también han identificado que los marcadores de activación de la mTORC1 no podría estar siempre correlacionada con la respuesta sintética de proteínas (Mitchell et al., 2013) o la adaptación crónica del ejercicio resistido (Phillips et al., 2013).
Señalización intracelular regulada por el ejercicio concurrente en el músculo esquelético
Se ha propuesto que la supresión mediada por la AMPK a la mTORC1 es el principal mecanismo por el que el ejercicio de edurance puede embotar las ganancias de fuerza después del ejercicio de resistido (Hamilton & Philp, 2013). De hecho, mientras que el estímulo de crecimiento proporcionado por el ejercicio de resistido inhibe la corriente del complejo mTORC1 represor TSC1-TSC2, el ejercicio edurance parece promover los efectos opuestos de una manera dependiente de la AMPK (Figura 2)
Sin embargo, aunque ambos procesos moleculares que controlan el anabólismo (mTORC1) y el catabólico (AMPK) coinciden con el efecto concurrente reportado en los seres humanos, también han informado algunos estudios resultados contradictorios, por lo que los eventos moleculares en el entrenamiento concurrente en la actualidad no está del todo claro.
Por ejemplo, el entrenamiento concurrente crónico se ha demostrado que induce efectos similares sobre la fuerza máxima y el área trasversal (CSA) como en el entrenamiento de fuerza, mientras que VO2max y el rendimiento físico mejoró a una semejante medida como con el entrenamiento de intervalo (de Souza et al., 2013).
Ejercicios de resistencia de intensidad moderada no parece afectar los efectos de la resistencia aguda ejercicio sobre mTORC1 y S6K fosforilación (Apro, Wang, Ponten, Blomstrand, y Sahlin, 2013) o la respuesta de la hipertrofia de corto plazo entrenamiento de la resistencia de una sola pierna (Lundberg, Fernández-Gonzalo, Gustafsson, y Tesch, 2013); sin embargo, ejercicio de intervalos de alta intensidad hace S6K1 romo fosforilación (Coffey et al., 2009). Además, la realización de ejercicio edurance antes del ejercicio resistido en un modelo simultáneo no tiene efectos perjudiciales en las señales anabólicas (Lundberg, Fernández-Gonzalo, Gustafsson, y Tesch, 2012) o de adaptación crecimiento a la formación de una sola pierna crónica (Lundberg et al., 2013). De hecho, al contrario del concepto de interferencias que hablamos anteriormente, el cual hay durante el entrenamiento concurrente, se ha mostrado mejora miofibrilar y mitocondrial (Cámara et al., 2014) como las tasas de síntesis de proteínas fraccionadas (Donges et al., 2012) obviamente en el estado alimentado a un grado mayor que si se hace solamente el ejercicio resistido. Estos datos, por lo tanto, indica que la respuesta aguda a nivel fisiológico y molecular al ejercicio concurrente son altamente dependientes sobre los detalles del entrenamiento a llevar a cabo, régimen, el modo de ejercicio y el tiempo empleado (agudo vs modelos crónicos). Como tal, contemporánea
Los estudios todavía no han identificado una unificación en el mecanismo que representa la interferencia inicial que hablamos anteriormente observada por Hickson. Teniendo en cuenta que se produce la interferencia después de 8 semanas de entrenamiento concurrente (Hickson, 1980; Ronnestad et al., 2012), los protocolos de entrenamiento de manera crónica combinado con estudios agudos específicos parecen representar el enfoque más apropiado para delinear los mecanismos moleculares que median la interferencia en la formación del músculo esquelético humano (Fyfe et al, 2014;. Hamilton & Philp, 2013).
Los métodos de nutrición para mejorar el rendimiento en el entrenamiento resistido y el rendimiento en edurance- implicaciones para entrenamiento concurrente
Mientras que el efecto de interferencia hace que el entrenamiento concurrente un desafío técnico, sino que también añade una complejidad adicional al considerar las necesidades nutricionales para un atleta que se entrena de manera concurrente. En términos generales, la restricción de energía en el músculo esquelético parece potenciar negativamrnte el medio ambiente de señalización entrenamiento de resistencia, mientras que un estado energético positivo, es decir un balance es óptimo podría maximizar el entorno anabólico post-ejercicio (Hamilton & Philp, 2013). La investigación detallada de los requerimientos nutricionales para el entrenamiento concurrente no se ha llevado a cabo, por lo que gran parte de nuestro el conocimiento acerca de las estrategias nutricionales que pueden influir el entrenamiento concurrente proviene de estudios asilados que analizan el impacto de las intervenciones nutricionales en el entrenamiento resistido o el entrenamiento edurance. Basado en estos estudios, la siguiente sección examinará estrategias nutricionales utilizadas para resaltar prácticas comunes, de diferentes modos que podrían ser adecuados para el atleta concurrente. Además, dada la necesidad de sintetizar nuevas proteínas musculares (por ejemplo, de generación fuerza miofibrilar y / o de producción de energía mitocondrial) a fin de facilitar la recuperación muscular y, finalmente, mejorar las adaptaciones crónicas al entrenamiento para el atleta concurrente, la siguiente sección leer aqui dejare las respuestas moleculares, y me centraré en gran medida en el papel que juega la alimentación en la remodelación del músculo esquelético.


