Resumen
El entrenamiento de la fuerza y de la resistencia produce adaptaciones ampliamente diversificadas, con muy poco solapado entre ellos. El entrenamiento de la fuerza típicamente produce aumentos en la masa muscular y fuerza muscular. En contraste, el entrenamiento de la resistencia induce aumentos en el consumo máximo de oxígeno y adaptaciones metabólicas que llevan a una capacidad de ejercicio mayor. En muchos deportes, una combinación del entrenamiento de la fuerza y de la resistencia es requerido para mejorar el rendimiento, pero en algunas situaciones cuando el entrenamiento de la fuerza y de la resistencia se realiza simultáneamente, una potencial interferencia en el desarrollo de la fuerza tiene lugar, haciendo tal combinación aparentemente incompatible. El fenómeno del entrenamiento concurrente, o simultáneamente el entrenamiento de la fuerza y resistencia, se describió primero en la literatura científica en 1980 por Robert C. Hickson, y aunque el trabajo que siguió dio evidencia a favor y en contra de él, el efecto de la interferencia parece sostenerse verdaderamente en situaciones específicas. A nivel molecular, allí parece estar una explicación para la interferencia del desarrollo de la fuerza durante el entrenamiento concurrente; está ahora claro que las formas diferentes de ejercicio inducen mecanismos de señalización intracelulares antagónicos que, a su vez, podría tener un impacto negativo en la respuesta adaptativa del músculo a esta forma de entrenamiento particular. Es decir, la activación de la AMPK por el ejercicio de resistencia puede inhibir la señalización a la maquinaria síntesis de proteínas inhibiendo la actividad de la mTOR y sus blancos-objetivos en la misma dirección. El propósito de esta revisión es describir el problema del entrenamiento concurrente de la fuerza y de la resistencia brevemente y examinar nuevos datos que resaltan mecanismos moleculares potenciales que pueden ayudar a explicar la inhibición del desarrollo de la fuerza cuando el entrenamiento de la fuerza y de la resistencia se realizan simultáneamente.

Las adaptaciones al ejercicio son muy dependientes del tipo de entrenamiento específico realizado (4,20,26,35). El entrenamiento de la resistencia, que representa un extremo de la actividad física, generalmente abarca duraciones de ejercicio de varios minutos a varias horas a distintas intensidades de ejercicio, aumentando la capacidad de sostener ejercicio repetitivo de alta intensidad, ejercicio de baja resistencia como pedalear, correr, y nadar. Esta mayor capacidad para rendir es principalmente cumplida a través de un aumento en el consumo máximo de oxígeno (O2máx) y una capacidad mayor del músculo esquelético para generar energía vía el metabolismo oxidativo sin mejoras en la fuerza muscular (4,20). El entrenamiento de la fuerza, que representa el otro extremo de la actividad física, abarca la actividad de corta duración a intensidades de ejercicio altas o máximas, aumenta la capacidad de realizar ejercicio de alta intensidad y alta resistencia de únicas o relativamente pocas repeticiones como el levantamiento de pesas olímpico, el powerlifting, y eventos de lanzamientos en el atletismo. El rendimiento mejorado relacionado con la fuerza es cumplido a través del aprendizaje neuromuscular y el mayor sincronismo del reclutamiento de fibras, hipertrofia de la célula muscular, y, posiblemente, hiperplasia, sin cambios en el O2máx o en la capacidad de generar el ATP vía metabolismo oxidativo (26,27,31). Dado tales modos contrastantes de ejercicio, y el hecho de que un número grande de actividades deportivas como la carrera de sprint (media distancia), rugby, fútbol, natación, y decatlón (entre muchos otros) parecen requerir combinaciones de ambos componentes de entrenamiento de la fuerza y de la resistencia para el rendimiento máximo, un modelo hipotético de un ‘continuum’ de fuerza-resistencia (CFR) puede definirse para ilustrar el rango de fuerza, resistencia, y las combinaciones metabólicas que el entrenamiento deben enfatizar para el mejor rendimiento (Fig. 1). Como consecuencia, el entrenamiento para muchos de estos deportes encontrará algunos problemas logísticos y algunas posibles limitaciones biológicas probablemente durante el curso del desarrollo del rendimiento.

En vista de las adaptaciones divergentes inducidas por los regímenes de entrenamiento de la fuerza y la resistencia, y las limitaciones potenciales observadas cuando se realizan ambas formas de ejercicio simultáneamente, la meta principal de la presente revisión es describir brevemente, en base a la evidencia existente, si el entrenamiento de la fuerza y la resistencia simultáneo resulta en rendimientos mejorados o disminuidos que cuando ocurren cualquier tipo de entrenamiento se ha realizado exclusivamente (por separado). Una segunda meta es delinear los mecanismos fisiológicos, bioquímicos, y moleculares potenciales asociados con las respuestas del músculo esquelético a las formas diferentes de ejercicio que pueden contribuir a la interferencia del desarrollo de la fuerza durante el entrenamiento concurrente.

Figura 1. El continuum de fuerza-resistencia (CFR) se describe en el contexto del rendimiento deportivo y su relación a la duración y al metabolismo energético. Desde un punto de vista energético-muscular y de la especificidad del entrenamiento, actividades de ejercicio de varios segundos hasta 1 minuto generalmente usa fuentes inmediatas de energía como el ATP, fosfato de creatina, y glucólisis, y requieren potencia y fuerza máximas. Las duraciones de ejercicio de varios minutos generalmente usan la glucólisis, oxidación de glucosa, y alguna oxidación de ácidos grasos que requieren el consumo de O2 cerca del máximo o máximo con distintos grados de fuerza. El ejercicio de largo duración de aproximadamente 20-30 minutos hasta varias horas de duración, usan principalmente la glucólisis aeróbica y la oxidación de ácidos grasos a tasas de trabajo submáximas tan cerca del O2máx como sean posibles, y parece requerir una cantidad pequeña de fuerza. Aunque el entrenamiento para los deportes de los extremos del CFR parece relativamente sencillo, una situación más complicada emerge cuando se diseñan los programas de entrenamiento para esos deportes que requieren combinaciones de fuerza y resistencia y una mezcla de fuentes generadoras de combustible.
LA INCOMPATIBILIDAD DEL ENTRENAMIENTO CONCURRENTE DE FUERZA Y RESISTENCIA: LOS EFECTOS SOBRE EL DESARROLLO DE LA FUERZA
El objetivo del estudio inicial que describe el fenómeno de entrenamiento concurrente era determinar cómo los individuos se adaptarían a una combinación de entrenamiento extenuante de fuerza de pesada resistencia y resistencia de alta intensidad comparado con las adaptaciones producidas ya sea o por el entrenamiento mismo de la fuerza o de la resistencia separadamente. Este estudio fue publicado por Robert C. Hickson en 1980 (19) e (intelectualmente) originado durante sus estudios de post-doctorado con el Dr. John Holloszy después de que un programa de carrera se había agregado a su régimen de entrenamiento de la fuerza corriente (Robert C. Hickson, comunicación personal, 1997).
Las siguientes consideraciones se tuvieron en cuenta en el plan experimental: a) ambos tipos de entrenamientos involucrarían los mismos grupos musculares; b) la respuesta a los programas de resistencia y de fuerza no se solaparían, es decir, no habría aumento en la fuerza con el entrenamiento de la resistencia y no habría aumento en el O2máx con el entrenamiento de la fuerza; y c) la magnitud de cambio en las variables de criterio (O2máx y fuerza) sería bastante suficiente para detectar cualquier respuesta divergente por los grupos. Había tres grupos de ejercicio: un grupo de fuerza (F) que se ejercitó 30 min·d-1, 5 días·semana-1 durante 10 semanas; un grupo de resistencia (R) que se ejercitó 40 min·d-1, 6 días·semana-1 durante 10 semanas; y un grupo de F-R que realizó los mismos regímenes de ejercicio diarios como el grupo de F y el grupos de R combinado. Para los programas de entrenamiento de la fuerza, todos los ejercicios se realizaron con el mayor peso posible. En tanto la fuerza aumentaba, un peso adicional continuamente se ajustaba a lo largo del entrenamiento para mantener la resistencia máxima para las repeticiones requeridas. Similarmente, para los programas de entrenamiento de la resistencia, en tanto la potencia de los sujetos aumentaba durante el entrenamiento, la tasa de trabajo del pedaleo era también aumentada si era necesario para acercarse al O2máx. El programa de carrera consistió en carrera continua tan rápido como fuera posible durante 30 min·d-1 en la primera semana, 35 min·d-1 durante la segunda semana, y 40 min·d-1 después de esa semana. En el presente estudio, el O2máx se usó como la mejor variable de criterio para establecer un efecto del entrenamiento de resistencia. El O2máx, cuando fue medido durante el pedaleo o la carrera en cinta ergométrica y expresado en términos absoluto (L·min-1) o relativo (mL·kg-1·min-1), aumentó a la misma magnitud (20-25%) en el grupo de R y en el grupo F-R. El O2máx aumentó ligeramente (4%) en el grupo F durante el pedaleo; de lo contrario, el entrenamiento de la fuerza no produjo ningún otro cambio significativo en el O2máx cuando era expresado en los valores absolutos o relativos. Como se esperaba, el entrenamiento de la resistencia no aumentó la fuerza significativamente. El entrenamiento de fuerza produjo aumentos en la fuerza de tal forma que en una base semanal era posible notar la mejora significativa en la sentadilla paralela a lo largo del programa de entrenamiento de 10 semanas. En contraste, el entrenamiento con pesas intenso combinado con un programa de entrenamiento de la resistencia produjo una mejora significativa en la fuerza durante las primeras 6-7 semanas, seguido por un período de nivelación, y luego, sorprendentemente, la fuerza disminuyó durante las últimas 2 semanas del programa de entrenamiento (Fig. 2). Estos resultados proveen la primera evidencia en indicar que en los límites superiores del desarrollo de la fuerza, el entrenamiento de la resistencia inhibe o interfiere con más aumentos en la fuerza. Estos resultados también indican que hay relación pequeña entre la adquisición de fuerza y la tasa de aumento en la potencia aeróbica.

Figura 2. Interferencia del desarrollo de la fuerza por el entrenamiento simultáneo de la fuerza y resistencia como originalmente se reportó en la literatura científica. Reproducido de Hickson, R. C. Interference of strength development by simultaneously training for strength and endurance. Eur. J. Appl. Physiol. Occup. Physiol. 45:255-263, 1980.
Estudios adicionales han confirmado el hallazgo de que el entrenamiento concurrente interfiere con el desarrollo de la fuerza. Por ejemplo, Dudley y Djamil (13) estudiaron la combinación del entrenamiento de la resistencia en ciclismo en intervalos de alta intensidad y entrenamiento isocinético de la fuerza de alta velocidad. En este estudio, el O2máx de pedaleo aumentó a la misma magnitud (~18%) en el grupo R y en el grupo F-R cuando era medido varias veces a lo largo del período de 7 semanas, pero las mejoras de la fuerza eran diferentes entre los grupos F y F-R. El grupo F tuvo aumentos en el torque máximo a 0.00-4.19 rad·s-1, mientras que el grupo F-R sólo tuvo una mejora significativa a 0.00, 0.24, y 1.68 rad·s-1, indicando que, en este caso, la interferencia en el desarrollo de la fuerza ocurrió en ritmos de velocidad alta pero no en velocidad baja de la producción de la fuerza. Más datos que demuestran la interferencia del desarrollo de la fuerza por el entrenamiento concurrente fueron obtenidos por Kraemer y cols. (22), quienes encontraron que combinando el entrenamiento de la fuerza y de la resistencia afectaban los aumentos de la fuerza inducidos por el entrenamiento en las áreas transversales de las fibras. Tal observación indica que la interferencia del desarrollo de la fuerza también puede ocurrir a nivel celular. Además, estos autores también encontraron que el entrenamiento concurrente sólo comprometió el desarrollo de la fuerza cuando ambos modos de ejercicio sometieron el mismo grupo muscular, haciendo pensar otra vez en un efecto local en lugar de uno sistémico.
Otras investigaciones han reportado poca inhibición del desarrollo de la fuerza por el entrenamiento concurrente de fuerza y resistencia. Por ejemplo, Sale y cols. (32) entrenaron dos grupos, una pierna en uno grupo completó un programa de fuerza y la otra pierna un programa de resistencia de fuerza. En el segundo grupo, una pierna fue entrenada en resistencia y la otra entrenó resistencia y fuerza. El entrenamiento de la resistencia consistió en 5 turnos de 3 minutos de pedalear a tasas de trabajo que requerían el 90-100% del O2máx, mientras que el entrenamiento de la fuerza consistió en seis series de 15-22 repeticiones en el press de piernas a una resistencia máxima por un total de 22 semanas. Todos los tipos de entrenamientos produjeron respuestas similares, incluyendo mayor fuerza, mayor O2máx, y mayor actividad de la citrato sintetasa del músculo vasto externo. En vista de la similitud de las respuestas a estos regímenes, es entendible que ninguna inhibición de la fuerza se observó por la combinación del entrenamiento, porque los regímenes de entrenamiento parecen haber sido más sinérgicos en lugar de antagónicos.
Después de estos estudios iniciales, varias otras investigaciones o favorecieron o discreparon con la interferencia del desarrollo de la fuerza durante el entrenamiento concurrente. Muchas de esas diferencias entre los estudios del entrenamiento concurrente provienen de varios temas metodológicos y de organización. En base a la evidencia existente hasta ahora, el efecto de la interferencia parece ser verdadero en situaciones específicas. Algunos de los resultados diferentes fueron postulados para ser relacionados a la selección la variable dependiente (indicadores del resultado), modalidad de los programas de entrenamiento, características de los sujetos (la edad, el sexo, el estado de entrenamiento), y duración del estudio (24). Es más, tales diferencias hacen comparaciones difíciles a través de estudios diferentes lo que complica la comprensión de las adaptaciones al entrenamiento concurrente.
POTENCIALES MECANISMOS RESPONSABLES PARA LA INTERFERENCIA DEL DESARROLLO DE LA FUERZA DURANTE EL ENTRENAMIENTO CONCURRENTE DE FUERZA Y RESISTENCIA
Con los años, se han propuesto varios mecanismos como factores limitantes para la adaptación óptima del músculo esquelético, y se han identificado como «responsables» de o contribuir a la inhibición del desarrollo de la fuerza durante el entrenamiento concurrente. Éstos incluyen componentes neurales, disponibilidad de sustrato como combustible, transformación del tipo de fibra, sobreentrenamiento, y alteraciones en la síntesis de proteínas (24).
Componente neural.
Dudley y Djamil (13) y Chromiak y Mulvaney (8) han discutido la posibilidad de que los factores neurales y el reclutamiento de las unidades motoras pueden tener un rol significativo en la restricción del desarrollo de la fuerza con el entrenamiento de la fuerza y de la resistencia. Sin embargo, ningún factor específico todavía se ha aislado para apoyar este mecanismo fuertemente, con la excepción de un estudio de Hakkinen y cols. (15) en el que los efectos del entrenamiento concurrente sobre tasas de desarrollo de la fuerza fueron postulados de haber sido una consecuencia de los componentes neurales y del músculo, porque este tipo de entrenamiento atenuó el desarrollo de la fuerza explosiva limitando la rápida activación neural voluntaria.
Bajo contenido de glucógeno.
Sucesivos turnos de fuerza o ejercicio de resistencia pueden producir crónicamente un bajo nivel de glucógeno muscular que podría retardar o podría dañar los rendimientos subsecuentes. El entrenamiento de la resistencia repetido en días consecutivos puede reducir los niveles de glucógeno muscular de reposo en el músculo (9), y el vaciamiento del glucógeno se ha demostrado que ocurre después del ejercicio con pesas (35). Una posible implicación de los niveles bajos del glucógeno sobre la adaptación muscular inducida por el entrenamiento concurrente se resalta por los resultados de Creer y cols. (10), quienes recientemente reportaron que bajos niveles de glucógeno muscular dañaban las respuestas de la señalización intracelular a un turno agudo de ejercicio con pesas. Por lo tanto, llevando a cabo un programa de entrenamiento que dice sesiones diarias o incluso sesiones dos veces por día, pueden dañar las respuestas al ejercicio, y la recuperación del ejercicio y/o rendimiento durante la ejecución de sesiones de entrenamiento subsecuentes, por consiguiente, reduciendo la magnitud de las adaptaciones del entrenamiento de la fuerza.
Transformación del tipo de fibra.
Los cambios en la composición de la fibra muscular, particularmente como función de las alteraciones de la isomiosina, han sido considerados previamente como posible mecanismo de inhibición del desarrollo de la fuerza, asociada al entrenamiento de la resistencia (8,14). La hipertrofia del músculo esquelético después del entrenamiento de la fuerza ocurre a una magnitud mayor en las fibras de contracción rápida que en las fibras de contracción lenta (16,34). El entrenamiento intenso de la resistencia se ha observado que reduce la velocidad de acortamiento máxima (Vmáx) de las fibras tipo II o las fibras de contracción rápida, y que cambia el conjunto de la fibra muscular de esqueleto cuando es medido por los cambios en el ATPasa de la miosina (25,33), lo que indica que una reducción en el número relativo de las fibras tipo II por el entrenamiento de la resistencia podrían jugar un mayor rol limitando el desarrollo de la fuerza durante el entrenamiento concurrente.
Sobrentrenamiento.
Dos revisiones previas (8,14) sobre el entrenamiento concurrente de fuerza y resistencia han considerado el término «sobreentrenamiento» para dar cuenta de la incapacidad para lograr ganancias óptimas de fuerza cuando el entrenamiento de la fuerza y de la resistencia se realiza. El sobreentrenamiento sostiene un término bastante pobremente definido a pesar de los recientes esfuerzos por los fisiólogos del ejercicio para identificar sus orígenes. El sobreentrenamiento es un desequilibrio entre el entrenamiento y la recuperación (23). En general, se caracteriza por una disminución en el rendimiento o por una falta de su mejora. En el primer estudio de entrenamiento de la fuerza y de la resistencia, la fuerza disminuyó en las semanas 9na y 10ma del entrenamiento concurrente (19). Porque los sujetos estuvieron entrenándose 80 min·d-1, un argumento podría hacerse que el marcado deterioro del desarrollo de la fuerza por el grupo F-R era el resultado del desarrollo de una fatiga residual. Sin embargo, éste no pudo haber sido el caso. El trabajo de resistencia por semana realizado en el cicloergómetro aumentaba a aproximadamente la misma tasa en el grupo R y en el grupo F-R, particularmente durante las semanas 9na y 10ma de entrenamiento, en un momento cuando las ganancias de fuerza en el grupo F-R estaban disminuyendo dramáticamente (Fig. 2). Así, los efectos en el grupo F-R sobre el desarrollo de la fuerza parecen ser selectivos para la respuesta del entrenamiento de la fuerza. Es más, los estudios de Dudley y Djamil (13) y de Hickson (19), así como otros estudios del entrenamiento concurrente, abarcaron protocolos algo diferentes de entrenamiento de la resistencia y de la fuerza (intensidad, duración, frecuencia, tipo de entrenamiento), incluyendo la secuencia de días cuando se realizaron cualquiera de los dos o ambos tipos de entrenamiento. En base a estas diferencias, es difícil de identificar el/los factor(es) que llevan a la inhibición de la fuerza por el sobreentrenamiento durante ambos tipos de entrenamiento.
Recambio proteico.
Generalmente se han encontrado que turnos agudos de ejercicios de resistencia reducen las tasas de síntesis de proteínas totales de músculos esqueléticos mixtos durante el ejercicio. Esta depresión es transitoria y puede llevar a una disminución temporal en la síntesis de proteínas dentro de varias horas después del ejercicio (5,12,29). La superposición de turnos de ejercicios de resistencia con el ejercicio con pesas puede producir respuestas adaptativas anormales en la síntesis de proteínas y, por lo tanto, una disminución en el rendimiento relacionado con la fuerza, en parte, debido a la falta de aumento en las áreas transversales de las fibras musculares (22). Cuando es realizado varias veces por semana, tal combinación del entrenamiento puede ser suficiente para deteriorar los mecanismos de la síntesis de proteínas involucrados con la adaptación normal a los turnos individuales de ejercicio de fuerza, alterando así las adaptaciones a largo plazo al entrenamiento y producir anormales ganancias de fuerza dependientes de lo muscular. Otra posibilidad, aunque hipotético, es que la síntesis de proteínas adaptativa que es el resultado de cualquier forma de ejercicio puede crear alguna clase de incompatibilidad celular en la que la célula muscular necesita decidir si crece o controla la síntesis de su maquinaria metabólica.
SEÑALIZACIÓN INTRACELULAR DURANTE EL ENTRENAMIENTO DE LA FUERZA Y DE LA RESISTENCIA: LECCIONES DE LA BIOLOGÍA MOLECULAR
Nosotros estamos ahora en una fase en que las tecnologías de campo tales como la bioquímica y la biología molecular pueden permitirles a los científicos del ejercicio explorar la biología de la adaptación del músculo esquelético inducida por el ejercicio en términos más mecanicistas. Estudios sobre la fosforilación de la proteína de las moléculas de la señalización intracelular han empezado a revelar procesos reguladores celulares específicos inducidos por las diferentes formas de ejercicio. Por ejemplo, el ejercicio con pesas agudo que, con el tiempo, puede producir hipertrofia muscular, induce la activación de una red de señalización asociado con el crecimiento. Experimentos en humanos y roedores demostraron que un único turno de ejercicio con pesas produce una mayor actividad de la fosfatidil inositol trifosfato kinasa (PI3k, phosphoinositide-3-dependent kinase) (18), de la proteina B kinasa (PKB, protein kinase B) (28), de la diana de la rapamicina mamífera (mTOR, Mammalian Target of Rapamycin) (3), y de la proteína ribosomal S6 kinasa 1 (S6k1, ribosomal protein S6 kinase 1) (1,21,28). La activación de tal red de señalización por el ejercicio con pesas agudo modula la síntesis de la proteína muscular tanto en los animales como en los humanos (18,11). La activación de la PI3-k lleva a un aumento en la actividad de la PKB y en la actividad de la mTOR, y a una inhibición subsecuente (fosforilación) de la proteína cap-dependiente 4E-BP1 (3,18), la que a su vez, inhibe la traducción del mRNA cap-dependiente y, de ahí, la síntesis de proteínas vía secuestro de la iniciación del factor eucariótico 4E (eIF4E). Un aumento en la actividad del eIF4E producirá tasas de síntesis de proteína muscular mayores. El ejercicio de resistencia, por otro lado, está asociado con mecanismos de señalización relacionados a las adaptaciones metabólicas, como la activación de la señalización de la proteína kinasa AMP-activada (AMPK). Una de las funciones principales de la AMPK es supervisar el estado de energía de la célula; por lo tanto, los procesos regulados por la AMPK parecen estar relacionados al mantenimiento de la homeostasis energética (17,37). La actividad de la AMPK se modula principalmente por los cambios en los niveles de los fosfatos de energía y por una disminución en la carga de energía de la célula muscular, esto es, un aumento en la proporción de ADP/ATP. Tales fluctuaciones en la regulación metabólica como las que ocurren durante el ejercicio (38,39) también pueden causar cambios en la expresión del gen y en el contenido de sustrato vía la señalización de la AMPK (17,37).
Es interesante observar que los recientes estudios han mostrado actividades antagónicas entre los mecanismos de la señalización anabólica inducidos por la red de PI3k/mTOR/PKB/S6k1/4E-BP1 y la señalización modulada por la energía por la AMPK. Más precisamente, la activación de la señalización de la AMPK por un agonista farmacológico reduce la síntesis de la proteína del músculo esquelético inhibiendo la señalización de la mTOR, probablemente vía activación del complejo de la esclerosis tuberoso (TSC). Bolster y cols. (2) han encontrado que una inyección de AMPK análogo AICAR (5-aminoimidazole-4-carboxamide 1-beta-d-ribonucleoside) no tenía efecto sobre la actividad de la [alpha]1 AMPK, pero aumentó la actividad de la [alpha]2 AMPK, un ~50%. La activación de la AMPK por el tratamiento de AICAR se correlacionó con una disminución del 45% en la síntesis de proteínas y fue asociada con una menor activación de la vía PKB/mTOR/S6K1. Esto también fue asociado con una inhibición reducida de la proteína eIF4E-dependiente (4E-BP1) y una reducción en el eIF4E asociado con el eIF4G. Como previamente se mencionó, un estímulo potente para el aumento en la actividad de la AMPK es un aumento en la proporción de ADP/ATP; este mecanismo puede ayudar a explicar las observaciones previas de Bylund-Fellenius y cols. (7), quienes demostraron que la actividad contráctil producía un aumento en la proporción de ADP/ATP que, a su vez, se correlacionó con una caída en las tasas de síntesis de proteína muscular. Estos resultados indican que la disminución en la síntesis de proteínas normalmente vista durante la actividad contráctil podría ser mediada en parte por un aumento la actividad de la AMPK y una disminución concomitante en la respuesta anabólica en la misma dirección de la señalización de la mTOR. En realidad, Thomson y Gordon (36) han hecho una observación interesante recientemente en animales viejos en donde la masa muscular se correlacionó negativamente con la actividad de la AMPK, implicando esta kinasa una vez más en la modulación negativa de la masa del músculo esquelético.
Otro mecanismo potencial para la inhibición de la síntesis de proteínas durante la actividad muscular puede estar en el paso de alargamiento (6). En un reciente estudio, Rose y cols. (30) descubrieron un aumento rápido en la fosforilación del alargamiento eucariótico factor 2 (eEF2) durante el ejercicio de pedaleo. En este estudio, los sujetos se ejercitaron aproximadamente al 67% del O2máx, y se obtuvieron biopsias musculares en reposo y después de 1, 10, 30, 60, y 90 minutos del ejercicio. El ejercicio causó un rápido (dentro de 1 minuto) aumento (de cinco a siete veces) en la fosforilación del eEF2 que persistió durante el período del ejercicio entero. Sorprendentemente, una disminución bastante pequeña en la actividad de la kinasa del eEF2 (eEF2k) fue descubierta, indicando que incluso con una disminución menor en la actividad del eEF2k, la kinasa activada restante pudo haber sido suficiente para inhibir y, por lo tanto, fosforilizar el eEF2, causando una disminución en la síntesis de proteínas. Los mecanismos por los que la activación del eEF2k por el ejercicio parecen ocurrir en una forma calcio-dependiente, porque el eEF2k del músculo ejercitado era potentemente activado por la calmodulina calcio-dependiente (Ca2+CaM) in vitro. Esto indica que la fosforilación más alta del eEF2 en el músculo esquelético activo puede mediarse por una activación alostérica de la kinasa del eEF2 por la Ca2+CaM.
RESUMEN Y DIRECCIONES DE LA INVESTIGACIÓN FUTURA
Aunque hipotético, es razonable asumir que la activación de la AMPK y la inhibición del eEF2 por el ejercicio de resistencia y/o sesiones de ejercicio demasiado frecuentes chocará con las respuestas al ejercicio con pesas afectando los aumentos inducidos por el entrenamiento en la síntesis adaptativa de proteínas, porque la activación/inhibición de estas proteínas de la señalización pueden alterar independientemente o colectivamente a la inducción de una respuesta anabólica. La consecuencia obvia de tal antagonismo será un impacto negativo en las adaptaciones del entrenamiento de la fuerza a largo plazo, quizás cambiando la respuesta hipertrófica. Por lo tanto, éste puede ser un mecanismo responsable para los efectos perjudiciales observados del entrenamiento concurrente de fuerza y resistencia en el desarrollo de la fuerza (Fig. 3).

Figura 3. Un modelo activo de las redes de la señalización intracelular mediando sobre las adaptaciones de músculo esquelético inducidas por el ejercicio. El ejercicio con pesas es capaz de inducir un aumento en la actividad de una red que comprende la señalización de PI3k/PKB/mTOR/S6K1/S6/4E-BP1 para modular tasas de síntesis de proteínas y crecimiento muscular. El ejercicio de resistencia está asociado con una señalización involucrada en la homeostasis metabólica, comprendiendo la señalización de la AMPK, entre otros. La activación de la AMPK por el ejercicio de resistencia o la actividad contráctil, puede inhibir la señalización de la mTOR vía el complejo TSC y puede suprimir la síntesis de proteína muscular inducida por el ejercicio. Además, los flujos de calcio resultantes de la actividad contráctil prolongada puede inhibir la síntesis de proteínas de una manera Ca2+M-dependiente, inhibiendo el alargamiento de la cadena péptida vía la activación del eEF2k y un aumento subsecuente en la fosforilación del eEF2. La AMPK también puede inhibir la síntesis de proteínas activando el eEF2k.
Uno de los argumentos contra el entrenamiento concurrente es que las adaptaciones que el cuerpo humano experimenta en respuesta a diferentes estímulos de entrenamiento de fuerza y resistencia realizado en el mismo día de entrenamiento. Están en diferentes extremos de un espectro de un ‘continium’ de fuerza-resistencia (ver figura 1), ‘confundiendo’ al cuerpo en cómo debería responder a menos que favorables adaptaciones. Aquí es donde aparece el fenómeno de la Interferencia donde las ganancias de fuerza se ven comprometidas cuando se entrena simultáneamente con la potencia aeróbica, por ejemplo (ver figura 2).
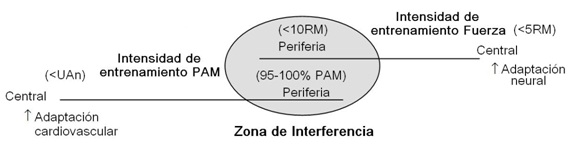
Figura 2. Contínuums de la intensidad y del lugar primario de adaptación del tanto el entrenamiento de la Potencia Aeróbica Máxima (PAM) como el entrenamiento de la fuerza, y el posible solapado cuando los 2 modos de entrenamiento son combinados. UAn: umbral anaeróbico; RM: máxima repetición;↑: aumento
nuestro foco de atención hoy está puesto sobre el entrenamiento concurrente y sus consecuencias sobre la salud de los individuos. Entonces, ¿qué incidencia tendría este tipo de entrenamiento sobre nuestra salud y el fitness en general? Como ejemplo, tenemos un estudio de J. Bentley, que utilizó sedentarios adultos durante 8 semanas de entrenamiento a 3 diferentes regímenes de ejercicio: entrenamiento aeróbico, entrenamiento de la fuerza y entrenamiento concurrente. El objetivo era observar cambios en el perfil de los lípidos en sangre y en la composición corporal de esos sujetos. El entrenamiento aeróbico consistía en un entrenamiento progresivo y periodizado de carrera en cinta ergométrica durante 16’ (1º semana) al 65% de la FCmáx, hasta 30’ al 80% (FCmáx) al final del período de entrenamiento. El entrenamiento de la fuerza consistía de un programa de entrenamiento progresivo usando pesos libres y máquinas en 4 ejercicios (press de banco, sentadilla, tirón de polea y curl de piernas, realizando 2×10 repeticiones al 50% de 1MR (1º semana) hasta llegar a 3×6 repeticiones al 80% de 1 MR al final del período, ajustando siempre las cargas conforme a las mejoras en la fuerza. En el entrenamiento concurrente se realizaban ambas formas de entrenamiento en el mismo día, separados por 15’ o 20’ entre cada tipo de ejercicio, realizando primeramente la sesión de fuerza para luego hacer la de resistencia.
Como resultado, los perfiles de las lipoproteínas de alta densidad (HDL) y de baja densidad (LDL) mejoraron con el entrenamiento de la resistencia y el entrenamiento concurrente, pero no con el entrenamiento de la fuerza. Los triglicéridos (TG) y el colesterol total (CT) mejoraron significativamente en los 3 regímenes de entrenamiento (ver figura 3).
La masa grasa total (MFT) disminuyó significativamente en el entrenamiento de la resistencia aeróbica y el entrenamiento concurrente, pero no en el entrenamiento de la fuerza, mientras que la masa libre de grasa (FFM) aumentó en el entrenamiento de la fuerza y en el entrenamiento concurrente, pero no en el entrenamiento aeróbico. Por lo tanto, se concluyó que el entrenamiento concurrente puede ser utilizado simultáneamente para mejorar el perfil de los lípidos en sangre y la composición corporal de sujetos previamente desentrenados.
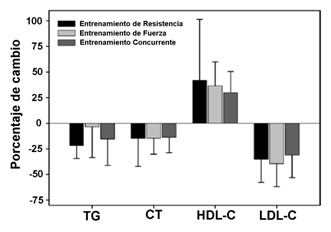
Figura 3. Porcentajes de cambios en el perfil de lípidos en sangre (Bentley, 2009)
En resumen, cuando el entrenamiento de la fuerza y de la resistencia se realiza simultáneamente, una interferencia potencial en el desarrollo de la fuerza puede ocurrir. Tal interferencia puede causarse por las alteraciones en los cambios de la síntesis de proteínas adaptativas, inducidos por el ejercicio de resistencia o por las sesiones de entrenamiento demasiado frecuentes, además de varios otros factores desconocidos. Las nuevas tecnologías están permitiéndonos entender los mecanismos moleculares involucrados en las adaptaciones del músculo esquelético inducidas por el ejercicio (por ejemplo, el análisis de la expresión gen del genoma amplio). Este conocimiento no sólo está mejorando la manera en que nosotros llevamos a cabo los programas de entrenamiento para los deportes y la rehabilitación, sino también es importante para el diseño correcto de estudios futuros en el ejercicio físico y la adaptación del músculo esquelético, particularmente aquellos que involucran la investigación de mecanismos de transducción de la señalización y de la expresión del gen.
Referencias
1. Baar, K., and K. Esser (1999). Phosphorylation of p70(S6k) correlates with increased skeletal muscle mass following resistance exercise. Am. J. Physiol. 276:C120-C127
2. Bolster, D. R., S. J. Crozier, S. R. Kimball, and L. S. Jefferson (2002). AMP-activated protein kinase suppresses protein synthesis in rat skeletal muscle through down-regulated mammalian target of rapamycin (mTOR) signaling. J. Biol. Chem. 277:23977-23980
3. Bolster, D. R., N. Kubica, S. J. Crozier, et al (2003). Immediate response of mammalian target of rapamycin (mTOR)-mediated signalling following acute resistance exercise in rat skeletal muscle. J. Physiol. 553:213-220
4. Booth, F. W., and K. M. Baldwin (1996). Muscle plasticity:energy demand and supply processes. In: Handbook of Physiology, Section 12: Exercise: Regulation and Integration of Multiple Systems, L. B. Rowell and J. T. Shepherd (Eds.). Bethesda, MD: American Physiological Society, pp. 1075-1123
5. Booth, F. W., and P. A. Watson (1985). Control of adaptations in protein levels in response to exercise. Fed. Proc. 44:2293-2300
6. Browne, G. J., and C. G. Proud (2002). Regulation of peptide-chain elongation in mammalian cells. Eur. J. Biochem. 269:5360-5368
7. Bylund-Fellenius, A. C., K. M. Ojamaa, K. E. Flaim, J. B. Li, S. J. Wassner, and L. S. Jefferson (1984). Protein synthesis versus energy state in contracting muscles of perfused rat hindlimb. Am. J. Physiol. 246:E297-E305
8. Chromiak, J. A., and D. R. Mulvaney (1990). The effects of combined strength and endurance training on strength development. J. Appl. Sports Sci. Res. 4:55-60
9. Costill, D. L., R. Bowers, G. Branam, and K. Sparks (1971). Muscle glycogen utilization during prolonged exercise on successive days. J. Appl. Physiol. 31:834-838
10. Creer, A., P. Gallagher, D. Slivka, B. Jemiolo, W. Fink, and S. Trappe (2005). Influence of muscle glycogen availability on ERK1/2 and Akt signaling after resistance exercise in human skeletal muscle. J. Appl. Physiol. 99:950-956
11. Cuthbertson, D. J., J. Babraj, K. Smith, et al (2006). Anabolic signaling and protein synthesis in human skeletal muscle after dynamic shortening or lengthening exercise. Am. J. Physiol. Endocrinol. Metab. 290:E731-E738
12. Dohm, G. L., G. J. Kasperek, E. B. Tapscott, and H. A. Barakat (1985). Protein metabolism during endurance exercise. Fed. Proc. 44:348-352
13. Dudley, G. A., and R. Djamil (1985). Incompatibility of endurance- and strength-training modes of exercise. J. Appl. Physiol. 59:1446-1451
14. Dudley, G. A., and S. J. Fleck (1987). Strength and endurance training. Are they mutually exclusive? . Sports Med. 4:79-85
15. Hakkinen, K., M. Alen, W. J. Kraemer, et al (2003). Neuromuscular adaptations during concurrent strength and endurance training versus strength training. Eur. J. Appl. Physiol. 89:42-52
16. Hakkinen, K., A. Pakarinen, M. Alen, H. Kauhanen, and P. V. Komi (1987). Relationships between training volume, physical performance capacity, and serum hormone concentrations during prolonged training in elite weight lifters. Int. J. Sports Med. 8(Suppl 1):61-65
17. Hardie, D. G., and K. Sakamoto (2006). AMPK: a key sensor of fuel and energy status in skeletal muscle. Physiology (Bethesda) 21:48-60
18. Hernandez, J. M., M. J. Fedele, and P. A. Farrell (2000). Time course evaluation of protein synthesis and glucose uptake after acute resistance exercise in rats. J. Appl. Physiol 1142-1149
19. Hickson, R. C (1980). Interference of strength development by simultaneously training for strength and endurance. Eur. J. Appl. Physiol. Occup. Physiol. 45:255-263
20. Holloszy, J. O., and F. W. Booth (1976). Biochemical adaptations to endurance exercise in muscle. Annu. Rev. Physiol. 38:273-291
21. Koopman, R., A. H. Zorenc, R. J. Gransier, D. Cameron-Smith, and L. J. van Loon (2006). The increase in S6K1 phosphorylation in human skeletal muscle following resistance exercise occurs mainly in type II muscle fibers. Am. J. Physiol. Endocrinol. Metab. 290:E1245-E1252
22. Kraemer, W. J., J. F. Patton, S. E. Gordon, et al (1995). Compatibility of high-intensity strength and endurance training on hormonal and skeletal muscle adaptations. J. Appl. Physiol. 78:976-989
23. Kuipers, H., and H. A. Keizer (1988). Overtraining in elite athletes. Review and directions for the future. Sports Med. 6:79-92
24. Leveritt, M., P. J. Abernethy, B. Barry, and P. A. Logan (2003). Concurrent strength and endurance training: the influence of dependent variable selection. J. Strength Cond. Res. 17:503-508
25. Luginbuhl, A. J., G. A. Dudley, and R. S. Staron (1984). Fiber type changes in rat skeletal muscle after intense interval training. Histochemistry 81:55-58
26. McDonagh, M. J. N., and C. T. M. Davies (1987). Adaptive responses of mammalian skeletal muscle to exercise with high loads. Eur. J. Appl. Physiol. Occup. Physiol. 56:178-198
27. Moritani, T., and H. A. deVries (1979). Neural factors versus hypertrophy in the time course of muscle strength gain. Am. J. Phys. Med. 58:115-130
28. Nader, G. A., and K. A. Esser (2001). Intracellular signaling specificity in skeletal muscle in response to different modes of exercise. J. Appl. Physiol. 90:1936-1942
29. Rennie, M. J., and K. D. Tipton (2000). Protein and amino acid metabolism during and after exercise and the effects of nutrition. Annu. Rev. Nutr. 20:457-483
30. Rose, A. J., C. Broholm, K. Kiillerich, et al (2005). Exercise rapidly increases eukaryotic elongation factor 2 phosphorylation in skeletal muscle of men. J. Physiol. 569:223-228
31. Sale, D. G (1988). Neural adaptation to resistance training. Med. Sci. Sports Exerc. 20:S135-S145
32. Sale, D. G., J. D. MacDougall, I. Jacobs, and S. Garner (1990). Interaction between concurrent strength and endurance training. J. Appl. Physiol. 68:260-270
33. Schantz, P., and J. Henriksson (1983). Increases in myofibrillar ATPase intermediate human skeletal muscle fibers in response to endurance training. Muscle Nerve 6:553-556
34. Staron, R. S., E. S. Malicky, M. J. Leonardi, J. E. Falkel, F. C. Hagerman, and G. A. Dudley (1990). Muscle hypertrophy and fast fiber type conversions in heavy resistance-trained women. Eur. J. Appl. Physiol. Occup. Physiol. 60:71-79
35. Tesch, P. A., E. B. Colliander, and P. Kaiser (1986). Muscle metabolism during intense, heavy-resistance exercise. Eur. J. Appl. Physiol. Occup. Physiol. 55:362-366
36. Thomson, D. M., and S. E. Gordon (2005). Diminished overload-induced hypertrophy in aged fast-twitch skeletal muscle is associated with AMPK hyperphosphorylation. J. Appl. Physiol. 98:557-564
37. Winder, W. W (2001). Energy-sensing and signaling by AMP-activated protein kinase in skeletal muscle. J. Appl. Physiol. 91:1017-1028
38. Wojtaszewski, J. F., C. MacDonald, J. N. Nielsen, et al (2003). Regulation of 5’AMP-activated protein kinase activity and substrate utilization in exercising human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 284:E813-E822
39. Yu, M., N. K. Stepto, A. V. Chibalin, et al (2003). Metabolic and mitogenic signal transduction in human skeletal muscle after intense cycling exercise. J. Physiol. 546:327-335
Cita
Cita Original
Nader GA. Concurrent strength and endurance training: from molecules to man. Med Sci Sports Exerc. 2006 Nov;38(11):1965-70.


